referido hecho anatómico y la mecánica cardíaca, coherencia siempre habida entre la forma y función de todo órgano, ha inducido al desarrollo de unos estudios experimentales según los cuales la disminución del volumen de las cavidades ventriculares, acompañada de un descenso de la base de los ventrículos, tiene lugar gracias a la contracción agonista del segmento descendente de la banda, previamente sometido a una elongación rectilínea, mientras el aumento del volumen ventricular, acompañado de un ascenso de la base de los ventrículos, acaece a consecuencia de la contracción antagonista del segmento ascendente de la banda, previamente sometido a una elongación curvilínea.
Palabras clave
SEGMENTO DESCENDENTE Y SEGMENTO ASCENDENTE
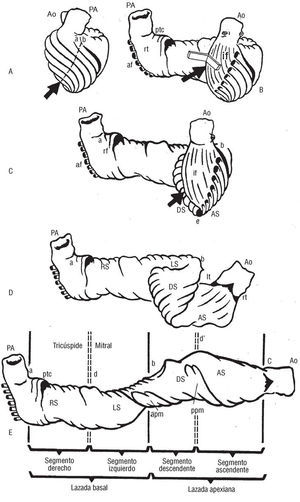
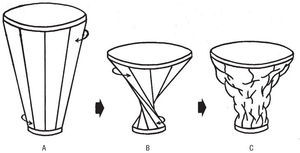
Las cavidades ventriculares vienen definidas por una banda miocárdica1 que describe dos vueltas de espiral en su discurrir desde la raíz de la arteria pulmonar hasta la raíz aórtica (fig. 1), configurando una helicoide. En la banda se pueden distinguir una lazada basal (fig. 2 a y b en el esquema E) y una lazada apexiana (fig. 2 b y c). A su vez, tanto en la lazada basal como en la apexiana se pueden diferenciar dos segmentos, el derecho (RS, a-d) y el izquierdo (LS, d-b), pertenecientes a la lazada basal, y el descendente (DS, b-d') y el ascendente (AS, d'-c) componentes de la lazada apexiana.

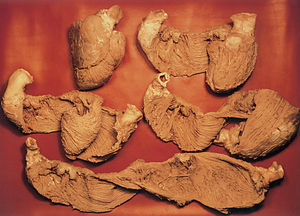
Fig. 1. Cinco corazones bovinos de similar tamaño muestran la secuencia de los cinco momentos más relevantes de la disección de la banda miocárdica ventricular que conforma las cavidades ventriculares al describir en el espacio, durante su trayectoria desde la raíz de la arteria pulmonar a la raíz aórtica, una helicoide con dos vueltas de espiral.

Fig. 2. Representación esquemática de la banda miocárdica ventricular.
HISTÉRESIS INTRÍNSECA E HISTÉRESIS EXTRÍNSECA
Con la palabra histéresis, de origen griego, se hacía referencia al tiempo transcurrido entre el momento de la aparición de una causa y su correspondiente efecto. Actualmente, sin embargo, se utiliza tal vocablo para designar el fenómeno por el cual la configuración de un objeto dado depende de su estado previo. La recuperación de la forma inicial de un trampolín, que en un momento dado queda combado al soportar en su extremo libre a un bañista, obviamente conduce a la rectilínea configuración habitualmente adoptada por el mismo cuando no es utilizado. La deformación y la subsiguiente reformación (en el sentido de volver a adquirir una determinada forma) del trampolín representan un ciclo de histéresis. Este concepto puede igualmente ser aplicado a la deformación y reformación de la esférica configuración de una pelota de goma, al alargamiento y subsiguiente acortamiento de un muelle de acero, etcétera.
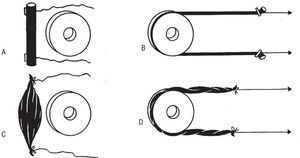
Una pieza de caucho alargada, como la representada en la figura 3A, al ser sometida a una tracción simultánea, ejercida desde sus dos extremos mediante sendas cuerdas fijadas en los mismos (B), queda deformada, con un patente aumento de su longitud y una disminución de su grosor, por adoptar una configuración curva al adaptarse a la superficie cilíndrica sobre la que se adosa. Al cesar la tracción, la pieza recupera su forma rectilínea inicial acortándose y engrosándose (A). En este caso, la recuperación de la forma inicial viene determinada por una fuerza endógena de naturaleza elástica, actualización de una energía potencial inherente, intrínseca, de la pieza en cuestión, que surge en determinadas circunstancias (p. ej., cuando el objeto elástico es incurvado como en la fig. 3B).

Fig. 3. Diferente respuesta de una pieza de caucho (A y B) y un músculo (C y D) a la tracción realizada por sus extremos y alrededor de un objeto cilíndrico. Mientras que la primera recupera espontáneamente la forma inicial, el músculo precisa de una estimulación eléctrica.
Pero algo muy distinto ocurre en el caso supuesto por un músculo (fig. 3C). Al efectuar una simultánea tracción de ambos extremos (D), mediante sendos cordeles fijados a los mismos, el músculo se alarga pero al cesar tal tracción no se recupera la forma inicial, que sólo puede lograrse mediante estimulación eléctrica. Con la llegada de este estímulo, la subsiguiente contracción determina entonces la recuperación de la configuración anterior (C). Evidentemente el estímulo eléctrico, desencadenante del fenómeno contráctil, es una energía de carácter exógeno procedente del entorno del músculo que, por tal razón, es ajena, extrínseca, al mismo. Cuando un individuo extiende un brazo, su bíceps queda alargado, al ser relajado isotónicamente, y no recupera espontáneamente su forma inicial acortándose por sí mismo gracias a una supuesta elasticidad; muy al contrario requiere la llegada de un estímulo eléctrico procedente del sistema nervioso central (del cerebro, acto voluntario, o de la médula espinal, acto reflejo), que por ende es ajeno a tal músculo, para que tenga lugar su contracción y el subsiguiente acortamiento.
Puede, por tanto, resumirse lo sucedido en los dos ejemplos más arriba referidos diciendo que para la de-formación representada por la elongación, tanto la de la pieza de caucho como la del músculo, se necesita la actuación de una fuerza, ajena al objeto, procedente del entorno (en estos dos ejemplos representada por la tracción efectuada con los cordeles (fig. 1B y D) mientras para la re-formación se requiere, en el caso de la pieza de caucho, la acción desencadenada por una energía intrínseca, propia del objeto (fuerza elástica inherente a la naturaleza del caucho) o una energía extrínseca, ajena al objeto (estímulo eléctrico procedente del sistema nervioso central), en el caso del músculo.
La distinta naturaleza de la energía desencadenante de la recuperación de la forma inicial permite, pues, hacer una distinción. En un objeto de caucho, o en un trampolín, un muelle, etc., se podría hablar de ciclos de histéresis intrínseca (histéresis propiamente dicha) mientras que en un músculo se debería hablar de histéresis extrínseca (falsa histéresis).
Pero en la recuperación de la forma inicial de los dos objetos de la figura 3, la pieza de caucho y el músculo, existe algo en común a pesar de la profunda diferencia existente entre las energías, elástica una y eléctrica otra, determinantes de la recuperación de la configuración original. En ambos objetos, esta recuperación da lugar, además de a un acortamiento, a un simultáneo envaramiento, enderezamiento o rectificación (A y C), de la curvatura (B y D) obligadamente antes adoptada a consecuencia de la tracción ejercida con los cordeles. La explicación de tal envaramiento representa el objetivo fundamental del presente trabajo.
AGONISMO Y ANTAGONISMO
Toda acción repetidamente llevada a cabo por un músculo dado implica, por principio, que vaya seguida de una acción opuesta desarrollada por otra fuerza ajena al mismo; en caso contrario, el resultado de cualquier actividad sólo podría ser alcanzado una sola vez. Se denomina, pues, músculo agonista a todo aquel cuya acción es neutralizada, anulada, por otro músculo denominado antagonista.
La reiteración de un determinado trabajo mecánico, propio de un ser vivo, viene condicionada por la actividad de un par muscular, agonista-antagonista, en el que cada uno de sus dos componentes con su actividad relaja isotónicamente al otro. No es adecuado pues afirmar que los músculos agonista y antagonista, de un determinado par muscular, son contrarios, impenitentes mutuos adversarios, como puede hacer pensar la etimología de la palabra con la que se designan (agonista, procedente del griego, significa combatiente). El músculo agonista no lucha contra el antagonista, toda vez que, por lo general, no realizan su actividad simultáneamente; muy al contrario lo que con su acción lleva a cabo el músculo agonista es favorecer al antagonista relajándolo isotónicamente, es decir, alargándolo, hecho por el cual se podría decir que lo prepara para que pueda volver a desarrollar su actividad, su trabajo. Y viceversa, exactamente lo mismo se puede afirmar del músculo antagonista. Todo músculo agonista y su correspondiente antagonista no luchan, pues, entre sí; muy al contrario, con su alternante actividad se favorecen mutuamente preparándose, uno al otro, para un eficaz desarrollo de su actividad.
CONTRACCIÓN AGONISTA Y CONTRACCIÓN ANTAGONISTA
Según lo ya referido cabe distinguir dos tipos de contracción: la agonista y la antagonista. La contracción agonista es aquella que provocando la flexión de determinada estructura (flexión de un brazo, por ejemplo, mediante la que un individuo aproxima hacia sí algo aprehendido con la mano) implica un acercamiento o aproximación, muchas veces con claros tintes de agresión a un objeto dado. La contracción antagonista, por el contrario, es aquella que provocando la extensión de determinada estructura (extensión de un brazo, por ejemplo, mediante la que un individuo aparta, lejos de sí, algo) implica un alejamiento o separación, no pocas veces sinónimo de evasión, de un objeto dado.
Pues bien, se puede decir que el corazón, cuya función se atiene estrictamente a tal doble normativa funcional, es el motor de la circulación de la sangre, mientras que los ventrículos son el motor del corazón, y que el motor de los ventrículos es la lazada apexiana, toda vez que la contracción agonista de su segmento descendente origina, mediante el acercamiento de la base al ápex, una disminución de volumen ventricular causa de la expulsión de la sangre a las grandes arterias, pulmonar y aorta, mientras la contracción antagonista de su segmento ascendente origina, mediante el alejamiento de la base del ápex, un aumento del volumen ventricular causa de la succión de la sangre de las aurículas.
ACORTAMIENTO Y ALARGAMIENTO
Esta doble función, expulsión y succión, se lleva a cabo mediante un respectivo alargamiento (fig. 4A) y acortamiento (B) del cono representado por la masa miocárdica ventricular2. Los ventrículos al acortarse disminuyen el volumen de sus cavidades mientras al alargarse lo aumentan, viniendo a sumarse, en la génesis de tales cambios volumétricos, un correspondiente engrosamiento y adelgazamiento de las paredes ventriculares.

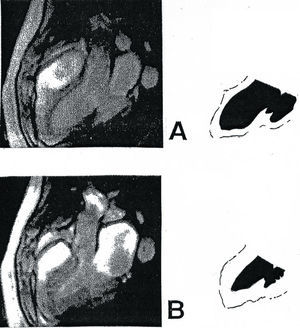
Fig. 4. Dos fotogramas de un film obtenido por resonancia magnética nuclear de un corazón humano normal latiendo, realizado por F. Herráiz en el Hospital Ruber Internacional de Madrid a petición del autor de este trabajo. A, imagen perteneciente a la fase de succión, que corresponde al final de la actividad contráctil ventricular (contracción del segmento ascendente) que es seguida por la fase de drenaje (ver fig. 10). B, imagen correspondiente a la fase de expulsión ventricular. Obsérvese, en ambos fotogramas, el cambio de volumen de las cavidades de los ventrículos así como la ostensible variación del espesor de sus paredes y el ascenso y descenso de la base ventricular.
Así, pues, el acortamiento y el alargamiento ventricular tienen lugar gracias a un correspondiente descenso y ascenso de la base ventricular, hecho un tanto paradójico dadas las relaciones anatómicas de la base, que la fijan a las grandes arterias y a las aurículas, y la libertad de movimientos del ápex que, a pesar de la total carencia de atadura alguna, permanece prácticamente inmóvil a lo largo de todo el ciclo cardíaco.
TORSIÓN Y DESTORSIÓN
Pero, a su vez, tales acortamiento y alargamiento de los ventrículos, causas respectivas de la disminución y aumento del volumen de sus cavidades, tienen lugar mediante una torsión, aquél, y una destorsión, éste, hecho fácilmente comprobable3-6 mediante resonancia magnética (tagging) y otros procedimientos experimentales.
La torsión de la masa ventricular determina un estrujamiento de la misma. Con su torsión los ventrículos son exprimidos, como una toalla mojada retorcida, y, consecuentemente, en principio cabría suponer que la destorsión podría representar la causa determinante de un aumento del volumen de las cavidades ventriculares. Sin embargo, esto no es así; en sí misma la destorsión no implica un aumento de aquel volumen.
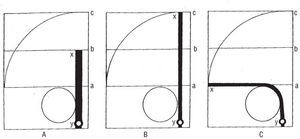
Como se aprecia en el símil de la figura 5A, la torsión (B) implica, necesariamente, un acortamiento del tronco de cono con una manifiesta disminución de su volumen, mientras la destorsión (C) puede tener lugar sin que ocurra necesariamente alargamiento alguno (como podría ser el representado en A) que determine un aumento de volumen. Es decir, durante la torsión (B) las dos bases del tronco de cono necesariamente se aproximan, mientras la destorsión no exige, en absoluto, separación alguna de las mismas, toda vez que tal destorsión puede efectuarse (C) sin que tenga lugar aquella separación. La torsión implica pues, por sí misma, aproximación de las bases, pero la destorsión no exige separación alguna; en cualquier caso sólo puede decirse que la permite.

Fig. 5. A: tronco de cono en el que, mediante la rotación opuesta de sus dos bases en el sentido marcado por las flechas, se efectúa una torsión, B, que, a su vez, mediante la rotación de ambas bases en sentido contrario, apuntado por las dos flechas, puede dar lugar a una destorsión tal como aparece en C (el vocablo destorsión, no reseñado en los diccionarios, ha sido introducido en el presente trabajo para expresar la acción contraria a la torsión). Evidentemente la torsión, B, exige, ineludiblemente, la aproximación de ambas bases del tronco de cono pero la destorsión no implica la separación de las mismas; tan sólo se puede decir que la permite.
Por tanto, ya que la destorsión por sí misma no puede implicar alargamiento de los ventrículos mediante una separación o alejamiento entre sí de la base ventricular y el ápex, el abordaje del problema planteado por el aumento de volumen de las cavidades ventriculares queda, pues, enfocado a determinar cuál es la fuerza que durante la destorsión de la masa ventricular origina el ascenso de la base de los ventrículos y, por ende, aquel aumento de volumen. Debe por tanto definirse tal fuerza precisando su origen, naturaleza (elástica o contráctil, no cabe otra posibilidad) y modo de actuación.
ELONGACIÓN RECTILÍNEA Y ELONGACIÓN CURVILÍNEA
En la figura 6A se representa, sobre una superficie plana, un objeto elástico en forma de tira alargada con uno de los extremos fijo (y) y el otro (x) libre. Todo cuerpo sólido capaz de acortarse en un momento dado, gracias a una energía endógena (histéresis auténtica, propia de objetos elásticos) o exógena (histéresis falsa, propia de los músculos, por ejemplo), con su retracción realiza un trabajo. Obviamente la retracción o acortamiento del objeto considerado sólo puede tener lugar si ha sido previamente alargado (significando en el caso del músculo una relajación isotónica y en el del objeto elástico una simple elongación), un alargamiento que puede ser rectilíneo, cuando el objeto en cuestión es sometido a un estiramiento en línea recta mediante tracciones opuestas aplicadas a sus extremos (fig. 6B), o curvilíneo, cuando además de la tracción se realiza una incurvación del mismo apoyándolo en algún objeto fijo (C). La diferenciación de estos dos hechos, los representados por una elongación rectilínea o curvilínea, aparentemente puede parecer banal, dado que lo conseguido en ambos casos viene a ser lo mismo, un alargamiento, pero, sin embargo, tiene una gran trascendencia, como será comprobado a continuación, para alcanzar el entendimiento de la función del corazón.

Fig. 6. Elongación rectilínea (B) y curvilínea (C).
El resultado a que da lugar un acortamiento (retracción, caso del cuerpo elástico, o contracción, caso del músculo) depende del tipo de elongación, rectilínea o curvilínea, previamente efectuado. En el caso de un objeto elástico, como el de la figura 6, la tracción rectilínea (B) determina que el objeto sea elongado en línea recta hasta una determinada longitud pasando su extremo libre (x) del nivel b, que presentaba en A, al nivel c ahora alcanzado en B. Si a continuación cesa la tracción, tal extremo libre (x) vuelve entonces al nivel b (A), recuperando la pieza su configuración inicial mediante un movimiento que tan sólo ofrece un único componente: el representado por un acortamiento.
Pero además de la tracción rectilínea se puede realizar otro tipo de tracción, la curvilínea (fig. 6C). Entonces el objeto elástico, en este caso doblado sobre un punto fijo más o menos circular, es incurvado, hecho por el cual el extremo libre (x) pasa del nivel b (que presentaba en A) al nivel a (ahora alcanzado en C). Si a continuación cesa la tracción, tal extremo libre (x) vuelve entonces al nivel b (el que presentaba en A) recuperando el objeto su configuración inicial mediante un movimiento en el que se pueden distinguir dos componentes (A): los representados por un acortamiento y por un simultáneo envaramiento (ver también la fig. 3A y C).
Este envaramiento o enderezamiento, que tiene lugar tras una tracción curvilínea, en el caso del objeto de la figura 6 origina, cuando cesa la tracción, un ascenso del extremo libre (x), que pasa del nivel a (en C) al nivel b (en A) gracias a la elasticidad del objeto en cuestión. De este modo se recupera la configuración inicial (A) completándose un ciclo de histéresis intrínseca. En el caso de un músculo, tal envaramiento sobreviene también tras efectuarse una tracción curvilínea del mismo (fig. 3D) aunque, en este caso, sea debido a la fuerza contráctil desencadenada por un estímulo eléctrico; se recupera de este modo la configuración inicial del músculo (fig. 3C) completándose un ciclo de histéresis extrínseca.
Así pues, estos dos ciclos, el de histéresis intrínseca del objeto de la figura 6, en el que la configuración inicial se recupera gracias a energía elástica (ver también la fig. 3B-A), y el de histéresis extrínseca del músculo, en el que la configuración inicial se recupera (fig. 3D-C) mediante energía eléctrica (estímulo), tienen algo en común; en ambos sobreviene aquel envaramiento mediante el que recobran su configuración inicial.
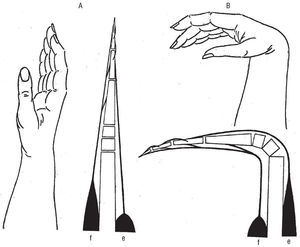
En el sistema musculoesquelético existen numerosos ejemplos de histéresis extrínseca, la desarrollada por todo par muscular agonista-antagonista. Un ejemplo muy representativo es el de los músculos flexores y extensores de los dedos de la mano. Cuando una mano extendida (fig. 7A) es flexionada (B), hecho que tiene lugar gracias a la contracción de los músculos flexores (agonistas, f) del antebrazo (hasta este momento alargados, extendidos rectilíneamente junto con sus tendones, tal como aparecen representados en A), los músculos extensores (antagonistas, e en B) se alargan quedando sus tendones incurvados sobre un punto de apoyo fijo, el representado por las epífisis distales del cúbito y radio. Si entonces sobreviene la contracción de tales músculos extensores, la mano adopta de nuevo su posición en extensión (A), hecho que, a su vez, implica la elongación (f en A) de los músculos flexores y sus tendones que, evidentemente, no podrían, por sí mismos, recuperar su configuración inicial (una imposibilidad igualmente aplicable a los músculos extensores cuando se contraen los flexores).
En consecuencia, de lo más arriba referido, cuando tras ser efectuada la flexión de una mano (fig. 7B) sobreviene la contracción de los músculos extensores, tal mano, basculando a nivel de la muñeca, asciende (A), hecho paradójico dado que tal ascenso tiene lugar gracias a un descenso, el representado por el acortamiento que implica la contracción de los músculos extensores (e en A). La explicación de tales fenómenos, por los que puede decirse que la maquinaria representada por el conjunto antebrazo-mano acusa un envaramiento, viene dada por una razón evidente: el distinto resultado a que da lugar una contracción agonista (f en B), como la de los músculos flexores cuando sus tendones, al estar la mano extendida (A) se hallan alargados tendidos en línea recta, y una contracción antagonista como es la de los músculos extensores (e en A) cuando sus tendones, al estar la mano flexionada (B), están también alargados pero curvados sobre un punto de apoyo fijo (las epífisis distales del cúbito y radio).

Fig. 7. Mediante una contracción antagonista, la de los músculos extensores del antebrazo (e), la mano adopta una posición en extensión, A. Mediante una contracción agonista, la de los músculos flexores del antebrazo (f), la mano adopta la posición en flexión, B. En este esquema B, el descenso de la mano, determinada por la contracción de los músculos flexores (f), implica una tracción curvilínea ejercida sobre los tendones de los músculos extensores (e) cuya incurvación, a nivel de la muñeca, es condición ineludible para que, después, tales músculos extensores puedan, con su contracción (e en esquema A), levantar la mano. Contracción agonista es sinónima de acortamiento, aproximación, agresión, apresamiento, mientras contracción antagonista es sinónima de alargamiento, separación, evasión, liberación.
La contracción antagonista (e en fig. 7A), determinante del envaramiento, representa una mera aplicación del principio mecánico de la palanca; en una palanca, que está descansando sobre un punto de apoyo central, el desplazamiento en un determinado sentido de un extremo de la misma, el de la potencia (músculos extensores en nuestro caso) implica un movimiento en sentido contrario del otro extremo, el de la resistencia (representada por la mano).
En el tan simple fenómeno mecánico representado por una contracción antagonista, que junto a un acortamiento del músculo implica un simultáneo envaramiento o enderezamiento de sus incurvadas fibras musculares o tendinosas, se basa, como se demostrará a continuación, la explicación del, a primera vista, paradójico aumento del volumen de las cavidades ventriculares determinado por una contracción muscular, la de las fibras miocárdicas constitutivas del segmento ascendente de la lazada apexiana.
EXPULSIÓN Y SUCCIÓN
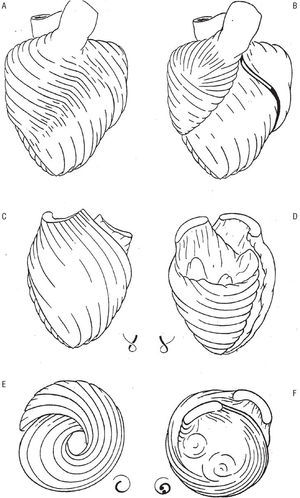
En la figura 8A aparece esquemáticamente reproducida la masa ventricular con las fibras aberrantes que, desde la cara anterior del ventrículo izquierdo, saltan sobre el surco interventricular anterior para recubrir el ventrículo derecho. Tras la eliminación primero de estas fibras (B) se reseca la lazada basal, al igual que la aorta, quedando aislada la lazada apexiana vista por delante (C), por detrás (D, cara diafragmática), por el ápex (E) y por la base (F).
La lazada apexiana, ha sido más esquemáticamente representada en la figura 9 (compárese el esquema A de esta figura con el esquema C de la fig. 8) a fin de facilitar la exposición de sus avatares funcionales.

Fig. 8. Cara anterior de la masa ventricular antes (A) y después (B) de eliminar las fibras aberrantes. Mediante la resección de la lazada basal, se aisla la lazada apexiana vista por la cara anterior (C), por la posterior o disfragmática (D), por el ápex (E) y por la base (F).

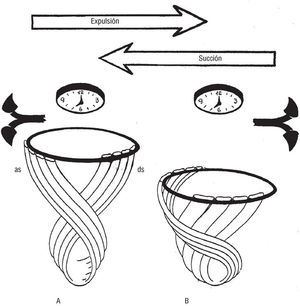
Fig. 9. Representación esquem ática de la lazada apexiana, en una vista anterior (compárese con la figura 8C), durante sus dos funciones. A) Succión ventricular. La contracción antagonista del segmento ascendente as (previamente incurvado en B) origina, a pesar del obligado acortamiento de sus fibras inherente a toda contracción, su envaramiento, acción que proyecta bruscamente hacia arriba a la base ventricular provocando el incremento del volumen de ambas cavidades ventriculares. Dicho de otro modo, en su condición de músculo antagonista el segmento ascendente origina, con la contracción de sus fibras (que, elongadas en B, recuperan ahora su configuración inicial rectificándose y engrosándose), un brusco ascenso de la base (expresado por la brusca caída de la presión intraventricular en las fases de presucción y succión de la fig. 10) junto con una rotación de la misma en sentido horario (flechas de la izquierda) y una elongación rectilínea del segmento descendente, ds, cuyas fibras, adelgazándose, recuperan ahora su grosor inicial. B) Expulsión ventricular. La contracción agonista de las fibras del segmento descendente origina un acortamiento de las mismas que tracciona hacia abajo a la base ventricular provocando el decremento del volumen de las cavidades ventriculares. Dicho de otro modo, en su condición de músculo agonista el segmento descendente (ds) origina simultáneamente, con su contracción (expresada en el esquema por el engrosamiento de sus fibras), un descenso de la base ventricular junto con una rotación de la misma en sentido antihorario (flechas de la derecha) y una elongación curvilínea del segmento ascendente (as).
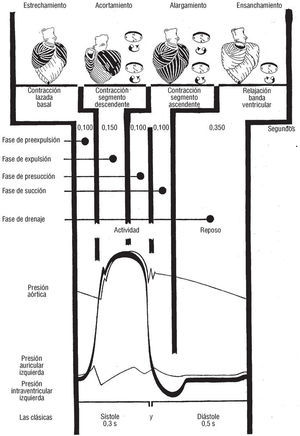
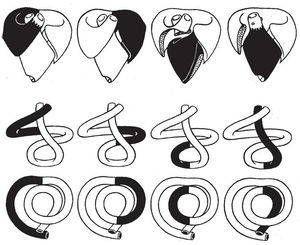
Tal como demuestra el estudio del análisis temporal secuencial de Fourier (análisis de fase o de primer armónico) de la ventriculografía isotópica7 y los resultados alcanzados mediante el emplazamiento intramiocárdico de cristales ultrasónicos8, la secuencia, de la entrada en actividad de las distintas regiones de los ventrículos, tiene lugar a lo largo de la banda miocárdica ventricular. Tras la contracción de la lazada basal (fig. 10, fase de preexpulsión) en la que entra primero en actividad el segmento derecho (fig. 11A) y después el izquierdo (B), sobreviene la contracción de la lazada apexiana que se inicia con la activación del segmento descendente (fig. 10, fase de expulsión; fig. 11C y fig. 9B) y finaliza con la del ascendente (fig. 10, fases de presucción y succión; fig. 11D y fig. 9A).
En la lazada apexiana (fig. 9) tiene lugar primero la activación del segmento descendente (esquema C de la figura 11), cuyas fibras, descendiendo desde la base ventricular hacia el ápex (ds, en la fig. 9A) en pronunciada configuración rectilínea, efectúan (B) una contracción agonista mediante la que tiene lugar un acortamiento (representado en el esquema de la fase de expulsión de la fig. 10) de la masa ventricular que, a su vez, implica:

Fig. 10. Cronología de las sucesivas fases del ciclo cardíaco.

Fig. 11. Secuencia de activación de los cuatro sucesivos segmentos, hecho experimentalmente comprobado mediante ventriculografía isotópica7.
1. Un descenso de la base de los ventrículos (figs. 9B y 5B).
2. una disminución del volumen ventricular (figs. 9B y 5B).
3. Un engrosamiento de las paredes ventriculares (fig. 4B).
4. Una elongación del segmento ascendente (fig. 9B).
5. Una rotación antihoraria de la base (fig. 9B).
6. Una torsión de la masa ventricular (fig. 9B).
Ocurre, entonces, la expulsión (fase de expulsión en la fig. 10) de la sangre habida en las cavidades ventriculares.
A continuación entra en actividad el segmento ascendente (esquema D de la figura 11) cuyas fibras, ascendiendo desde el ápex a la base ventricular (fig. 9B) en pronunciada configuración curvilínea, efectúan (A) una contracción antagonista mediante la que, al envararse, tiene lugar un alargamiento (fases de presucción y succión en la fig. 10) de la masa ventricular que, a su vez, implica:
1. Un ascenso de la base de los ventrículos (figs. 9A y 5A).
2. Un aumento del volumen ventricular (figs. 9A y 5A).
3. Un adelgazamiento de las paredes ventriculares (fig. 4A).
4. Una elongación del segmento descendente (ds en fig. 9A).
5. Una rotación horaria de la base (fig. 9A).
6. Una destorsión de la masa ventricular (fig. 9A).
Acaece, entonces, la succión de la sangre habida en las cavidades auriculares.
CONCLUSIONES
1. El aumento de volumen de las cavidades ventriculares es exclusivamente debido a una contracción.
2. La contracción, causa del aumento del volumen de las cavidades ventriculares, la realizan las fibras del segmento ascendente de la lazada apexiana.
3. La contracción del segmento ascendente de la lazada apexiana es una contracción antagonista que implica una separación o alejamiento de la base ventricular y el ápex.
4. La contracción antagonista del segmento ascendente requiere, como todas las de este género, un punto de apoyo.
5. El punto de apoyo del que se sirve el segmento ascendente para, mediante su contracción antagonista, alargar las cavidades de los ventrículos es el representado por el rígido núcleo constituido por la cavidad ventricular izquierda cuando alcanza su mínimo tamaño al estar tan sólo ocupada por el volumen residual. Tal papel de la cavidad ventricular izquierda, en la función diastólica del corazón, le confiere un protagonismo que refrenda su carácter de ventrículo frente al representado por la cavidad ventricular derecha, mera dehiscencia de la pared ventricular9.
6. La contracción antagonista del segmento ascendente tan sólo supone una mera aplicación del principio mecánico de la palanca.
7. Las dos espirales, descritas por la banda miocárdica ventricular en su recorrido desde la arteria pulmonar a la aorta (esquemas de abajo en la fig. 11; ver también las figs. 1 y 2), implican que los ventrículos funcionen como la musculatura circular (por ejemplo la de las paredes intestinales, vasos, bronquios, esfínteres, etc.) cuya contracción tiene lugar sobre un punto de apoyo móvil, el contenido de la cavidad tubular que tal musculatura circular delimita. Los alimentos en el tubo intestinal, la sangre en los vasos sanguíneos, el aire en los bronquios, etc., representan un punto de apoyo móvil, al igual que la sangre habida en las cavidades ventriculares, impelido a desplazarse cuando tiene lugar la contracción de la correspondiente musculatura circular. El corazón, cuya singular doble función se desarrolla con la más sutil, simple y eficiente mecánica que pueda ser llevada a cabo por fenómeno contráctil alguno, no requiere puntos de apoyo extrínsecos (como los propios de la musculatura esquelética perennemente anclada en rígidos huesos) para cumplir con su cometido; apoyado en el incompresible volumen residual, albergado por la cavidad ventricular izquierda, se basta a sí mismo para llevar a cabo consecutivamente acciones tan dispares como las de expulsar y succionar sangre.
Correspondencia: Dr. F. Torrent Guasp. La Plana, 63. 03700 Dénia. Alicante. Correo electrónico: ftorrentg@meditex.es