La teoría del funcionamiento del corazón de Torrent Guasp se basa en la agrupación de las fibras musculares que constituyen el miocardio ventricular en una banda continua que se enrolla en una doble hélice1,2.
Ni la observación directa o con instrumentos ópticos ni la palpación del órgano permiten identificar partes de la banda. La disección realizada por Torrent Guasp y quienes aprendieron de él reproduce en todos los corazones la misma estructura, y esta reproducibilidad es el único dato objetivable de la existencia de la banda como estructura anatómica aportado inicialmente.
Las técnicas de disección empleadas por muchos cardiólogos e investigadores, disección a punta de dedo en corazones previamente hervidos, no identifican planos de separación preexistentes y el resultado final es un artefacto que resulta similar a lo que ocurre cuando se reproducen obras de artesanía, se imita un modelo preconcebido y es la voluntad del autor lo que hace que las piezas sean iguales.
En general, para los fisiólogos y los biofísicos, la organización del miocardio en una banda con dos helicoides facilitaba mucho la comprensión de la mecánica cardiaca y su eficacia para generar la presión3,4. Muchas de las pruebas que se han aportado para demostrar que la banda es el sustrato anatómico real del corazón se derivan de esta «conveniencia fisiológica». La interdependencia entre estructura y función es máxima en el corazón y difícilmente se puede explicar el rendimiento cardiaco sin una estructura adecuada o aceptar una anatomía no compleja para el desarrollo competente de la función, aunque esto conlleva grandes dificultades en su estudio5–9. El artículo publicado en Revista Española de Cardiología por Poveda et al10 permite visualizar imágenes compatibles con trazados de fibras miocárdicas in situ con la propia forma y la complejidad que tiene la banda disecada una vez ordenada y «montada» en posición adecuada. Pero se trata de imágenes instantáneas obtenidas mediante un programa de ordenador que maneja señales de resonancia magnética (RM) procedentes de la conductividad de moléculas de agua entre las fibras miocárdicas de corazones de perro ex vivo. Se podría concluir que son la sombra de las fibras y unas piezas anatómicas muy sofisticadas, aunque lo más interesante es lo sugestivo de las imágenes.
¿QUÉ SON LAS IMÁGENES DE RESONANCIA MAGNÉTICA DEL TENSOR DE DIFUSIÓN DEL CORAZÓN?El tensor de difusión estudia la difusividad de las moléculas del agua. La información sobre el trazado de las fibras miocárdicas deriva del principio según el cual la orientación principal de las microestructuras del miocardio es paralela a la dirección de la máxima difusión de las moléculas de agua, cuya señal sufre una atenuación en presencia de un gradiente magnético11. De este modo, la RM proporciona en cada píxel de miocardio tres vectores ortogonales (eigenvectores) y sus magnitudes (eigenvalores). Mediante cálculos y modelos matemáticos, se obtienen tres parámetros fundamentales para definir las fibras miocárdicas: fracción de anisotropía, difusividad media y ángulo de hélice.
La difusividad media se calcula como un promedio de tres eigenvectores y sus cambios reflejan cambios patológicos que modifican la redistribución del volumen extracelular o intracelular.
La fracción de anisotropía es el grado de desviación que representa un elipsoide con respecto a una esfera (isotropía), se calcula con la desviación estándar de los tres eigenvectores del tensor normalizado por la magnitud de estos. Este parámetro mide la variabilidad en la movilidad del agua en diferentes direcciones y disminuye si se desorganiza la estructura tisular. Está relacionado con la arquitectura tisular y es un índice de integridad tisular. Cuando su valor es 0, indica que la difusión es isotrópica (en todas las direcciones), y cuando es 1, la difusión es anisótropica y ocurre sólo en una dirección.
La orientación de las fibras de cada pixel se define como el primer eigenvector del tensor, y el ángulo de hélice de la fibra es el ángulo entre el vector de la fibra local y el vector circunferencial local (el vector circunferencial es perpendicular tanto al eje largo del ventrículo izquierdo como al vector radial local) y representa la arquitectura de la fibra.
Las imágenes de RM de tensor de difusión caracterizan el tejido y nos permiten conocer la arquitectura de la fibra. Inicialmente se utilizaron en la cirugía del cerebro para seleccionar el tejido a extirpar, y en cuanto al corazón, hay algún trabajo publicado para reconocer el tejido afectado por el infarto de miocardio12, pero resulta muy difícil obtener imágenes validas por el movimiento del corazón y el de la respiración13.
Las imágenes que se presentan en el artículo de Poveda et al10 proceden de corazones de perro inmersos en una sustancia idónea por la baja sensibilidad al magnetismo. Los datos experimentales originales proceden de una base de datos de RM cardiacas. Los autores comparan las imágenes de RM del tensor de difusión con el trazado y la estructura de las diversas porciones de la banda plegada en su posición anatómica de un molde de goma que realizó Torrent Guasp. Las diversas partes de la banda coinciden en posición en el miocardio y en la dirección que trazan. Las imágenes no simplificadas de las figuras 2 y 3 del artículo de Poveda et al10 reproducen la imagen de la banda enrollada en su posición anatómica; por transparencia, se puede apreciar en el mesocardio el segmento descendente conformando el tabique y el ápex. También es sorprendente en las imágenes simplificadas el giro que se produce al inicio del segmento descendente apical y cómo se introduce en el subendocardio para formar el septo y el ápex.
Estas imágenes de RM se han obtenido en corazones explantados sin ninguna referencia posible a las fases del ciclo cardiaco. La gran pregunta es si en las imágenes del tensor de difusión puede haber información sobre la dinámica cardiaca. Concretamente, si la orientación de los vectores presentes en las figuras 1A y 1B del artículo de Poveda et al10 significa que la difusión del agua se realiza preferentemente o solamente en una dirección y que esta dirección está relacionada con la actividad mecánica del corazón. Los autores interpretan que la difusión del agua se puede producir en ambas direcciones, y no que la diferente orientación de los vectores se correspondiera con una u otra de las espiras, ya que en el doble helicoide una espira está dentro de la otra y la dirección de contracción puede corresponder a espiras diferentes según el ángulo de corte.
EVIDENCIAS FUNCIONALES DE LA EXISTENCIA DE UNA ESTRUCTURA EN BANDA ENROLLADAAlgunos investigadores han tratado de aportar evidencias funcionales que serían consecuencia obligada de la existencia y el funcionamiento de la banda3,5,8. Torrent Guasp interpretó que la onda de contracción recorrería la banda desde la zona subpulmonar donde localizaba el inicio hasta la raíz aórtica que constituía el final1,2. Así, diversos investigadores con distintos medios trataron de evidenciar esta cadencia de contracción. En nuestro laboratorio, utilizando la soniometría se pudo establecer esta cadencia midiendo el inicio de contracción en cuatro puntos del miocardio ventricular identificables con otros tantos puntos del recorrido de la banda muscular: a) epicardio de la zona subpulmonar (identifica el segmento derecho de la lazada basal); b) pared lateral del ventrículo izquierdo (identifica el segmento izquierdo de la lazada basal); c) mesocardio de la zona anterior del ápex ventricular izquierdo (identifica el segmento descendente de la lazada apexiana), y d) epicardio de la zona subaórtica (identifica el segmento ascendente de la lazada apexiana).
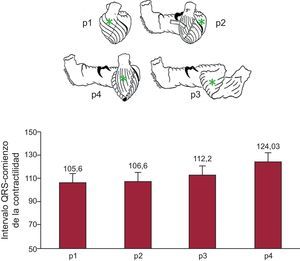
Se puede apreciar en la figura 1 los tiempos transcurridos entre el comienzo del complejo QRS y el comienzo de la contracción en los cuatro puntos descritos, recogidos mediante pares de cristales ultrasónicos implantados. Este progreso de la contracción a lo largo de la banda supone un paso cualitativo añadido; la estructura queda ligada a una dirección determinada de la contracción, con lo que la disposición de la banda y esa forma de contraerse son las encargadas de la función cardiaca.
. En la parte superior se puede ver la banda miocárdica desenrollada según Torrent Guasp. En ella están marcados los puntos (p) en los que se han implantado pares de cristales ultrasónicos siguiendo la dirección de las fibras. En la parte inferior, se representan en columnas los valores de media ± error típico de los tiempos de inicio de la contracción en los puntos señalados. La zona subpulmonar y la pared libre del ventrículo izquierdo son las zonas de contracción más precoz, seguidos del mesocardio de la cara anterior del ápex correspondiente al segmento descendente en la banda. Finalmente, se contrae el epicardio de la cara anterior del ventrículo izquierdo correspondiente al segmento ascendente de la lazada apexiana (datos propios no publicados).")
Experimentos en cerdos con tórax abierto (n = 32). En la parte superior se puede ver la banda miocárdica desenrollada según Torrent Guasp. En ella están marcados los puntos (p) en los que se han implantado pares de cristales ultrasónicos siguiendo la dirección de las fibras. En la parte inferior, se representan en columnas los valores de media ± error típico de los tiempos de inicio de la contracción en los puntos señalados. La zona subpulmonar y la pared libre del ventrículo izquierdo son las zonas de contracción más precoz, seguidos del mesocardio de la cara anterior del ápex correspondiente al segmento descendente en la banda. Finalmente, se contrae el epicardio de la cara anterior del ventrículo izquierdo correspondiente al segmento ascendente de la lazada apexiana (datos propios no publicados).
Las propiedades funcionales de las fibras miocárdicas podrían resumirse en: a) su contracción longitudinal que da lugar a un incremento en el grosor perpendicular al proceso de acortamiento; b) la activación eléctrica de una fibra se trasmite a las vecinas siguiendo la longitud de la banda, y c) hay fibras miocárdicas que conducen los estímulos más rápidamente, son las del tejido específico de conducción distribuido por el subendocardio y conectado directamente al sistema de conducción auriculoventricular. Esta última propiedad es la base de los mapas de activación eléctrica inicial del corazón, que difícilmente encaja con una cadencia de contracción a lo largo de la banda como la que hemos descrito previamente.
Son esenciales la dirección de las fibras y su agrupación en la banda. La dirección espacial de la contracción motivada por la propia arquitectura de la banda condiciona la eficacia del trabajo y la complejidad de su interpretación. Imaginemos la contracción lineal de un músculo esquelético en el que el único parámetro es contracción, ya que su cambio a grosor condiciona la función. Aquí el grosor reduce la cavidad, por lo que es un parámetro activo-eficaz. Dado que los puntos inicial y final de la banda están fijos, la reducción del volumen de la cavidad ventricular se sustancia en un incremento en el grosor de las paredes, además del efecto que tiene la contracción del segmento descendente de la lazada apexiana. El hecho de que la banda esté constituida por múltiples segmentos contráctiles uno a continuación del otro, por encima y por bajo de otros, supone que la dirección de contracción dentro del helicoide para cada porción es variable a lo largo del proceso y que los segmentos vecinos, sobre todo los que están en contracción, son los puntos de apoyo para los segmentos que inician su contracción.
La disposición helicoidal de la banda ya presupone movimientos de torsión con deslizamiento entre los dos helicoides sobre el eje longitudinal, que produce un efecto de tornillo y acerca y aleja del ápex la base ventricular.
En resumen, la activación y contracción sucesiva de los segmentos derecho e izquierdo de la lazada basal reducen transversalmente (fig. 1, puntos 1 y 2) y fijan la base del ventrículo izquierdo, la contracción del segmento descendente de la lazada apical (fig. 1, punto 3) aproxima atornillando la base al ápex, reduce longitudinalmente la cavidad ventricular y empuja la sangre hacia la cámara de salida del ventrículo izquierdo. La contracción precoz e independiente del subendocardio del ventrículo izquierdo genera una cavidad apexiana con una cierta morfología y suficiente rigidez para facilitar el efecto embolada. Probablemente para cuando se contraiga el segmento ascendente de la lazada apexiana (fig. 1, punto 4) y se inicie la diástole el subendocardio haya dejado de estar en contracción.
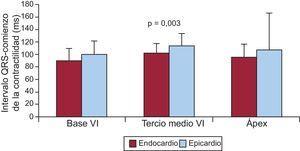
Esto supone una explicación a la aparente contradicción entre los mapas de activación eléctrica y la teoría del funcionamiento del corazón de Torrent Guasp y está basada en las medidas que realizamos comparando los tiempos de contracción del subendocardio y el epicardio en perros (fig. 2). Otros autores también han descrito este fenómeno de contracción precoz del endocardio14,15.
. Se representan en columnas los valores de media ± desviación estándar de los tiempos de inicio de la contracción en tres puntos epicárdicos escogidos al azar y sus correspondientes puntos en el endocardio. En todas las ocasiones la contracción fue significativamente más precoz en los puntos del endocardio (datos propios no publicados). VI: ventrículo izquierdo.")
Experimentos en cerdos con tórax abierto (n = 8). Se representan en columnas los valores de media ± desviación estándar de los tiempos de inicio de la contracción en tres puntos epicárdicos escogidos al azar y sus correspondientes puntos en el endocardio. En todas las ocasiones la contracción fue significativamente más precoz en los puntos del endocardio (datos propios no publicados). VI: ventrículo izquierdo.
Se acepta que la diástole ventricular es un proceso activo ligado a la contracción muscular7,9. La contracción del segmento ascendente de la lazada apexiana desenrosca la base, la separa del ápex y produce un efecto de succión que genera una presión negativa en la cámara ventricular e inicia el flujo transmitral; diversos autores han aportado experiencias que demuestran dicho efecto de succión16 y cómo este desaparece al bloquear la contracción del último segmento de la banda muscular8.
CONCLUSIONESEl funcionamiento del corazón es muy complejo. Para conocerlo, se abrieron nuevas perspectivas con la disección de su pared en la banda miocárdica. En este número se incluyen imágenes procedentes del tensor de difusión de RM que reproducen la banda descrita por Torrent Guasp ordenada y plegada en su posición anatómica. La existencia real de la banda ha facilitado la comprensión de la función cardiaca, pero por cada respuesta que se obtiene aparecen muchas más preguntas. Así, si la contracción se inicia en la zona subpulmonar (además del subendocardio que tiene la red de Purkinje), ¿cómo le llega la activación eléctrica? Son necesarias nuevas investigaciones, otros métodos y mentalidades capaces de romper con conocimientos preestablecidos.
CONFLICTO DE INTERESESNinguno.
Los autores agradecen los conocimientos aportados sobre el tensor de difusión de RM a la Dra. Alicia Maceira González.